Rozkład glukozy do pirogronianu to jeden z najważniejszych procesów w biologii komórki. Ja porządkuję go przez trzy pytania: skąd bierze się energia, co dzieje się z pirogronianem i kiedy komórka przyspiesza albo hamuje cały szlak. Dzięki temu łatwiej zrozumieć nie tylko sam mechanizm, ale też to, dlaczego jest on tak ważny w mięśniach, mózgu, erytrocytach i wątrobie.
Najważniejsze fakty, które warto zapamiętać od razu
- Szlak zachodzi w cytozolu i nie wymaga tlenu, ale dalszy los pirogronianu zależy już od warunków w komórce.
- Na jedną cząsteczkę glukozy komórka zużywa 2 ATP, a wytwarza 4 ATP i 2 NADH, więc bilans netto wynosi 2 ATP.
- Najważniejszym punktem regulacji jest fosfofruktokinaza-1, czyli enzym, który bardzo dobrze pokazuje stan energetyczny komórki.
- W erytrocytach końcowym produktem jest mleczan, bo nie mają mitochondriów.
- W tkankach tlenowych pirogronian zwykle trafia do mitochondrium i dalej zasila cykl Krebsa oraz fosforylację oksydacyjną.
Na czym polega szlak glikolityczny i kiedy komórka z niego korzysta
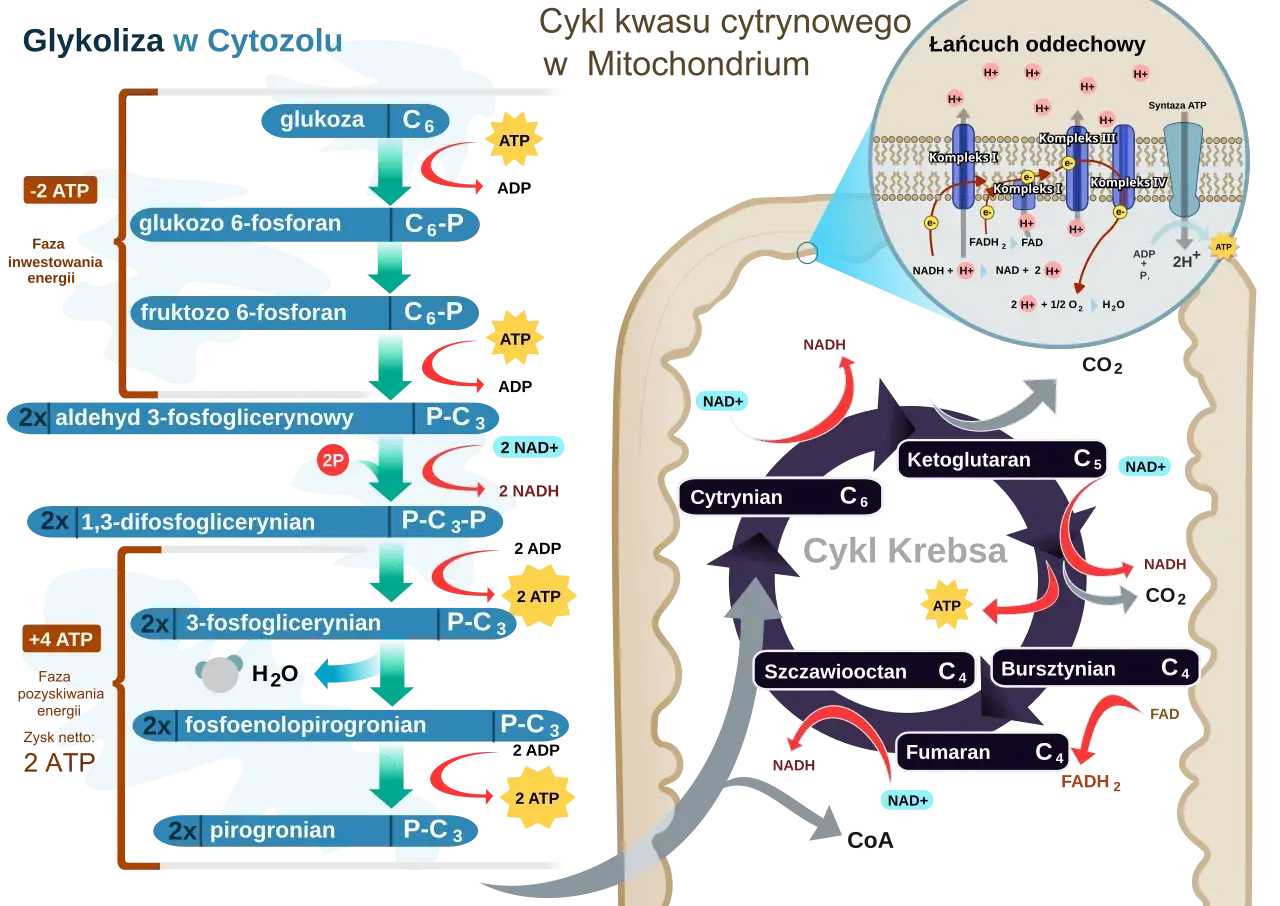
To podstawowy szlak kataboliczny, w którym komórka rozkłada glukozę do dwóch cząsteczek pirogronianu. W praktyce oznacza to szybkie uzyskanie energii, nawet wtedy, gdy dostęp tlenu jest ograniczony albo gdy komórka nie ma jeszcze sprawnego „zaplecza” mitochondrialnego. Właśnie dlatego ten proces jest tak uniwersalny: działa w wielu tkankach, a w wielu organizmach jest jednym z pierwszych i najbardziej konserwatywnych etapów pozyskiwania energii.
Patrzę na niego jak na mechanizm startowy. Nie daje maksymalnej ilości ATP z cząsteczki glukozy, ale jest szybki, elastyczny i działa w szerokim zakresie warunków. To również pierwszy etap oddychania komórkowego, więc jego znaczenie wykracza daleko poza samą produkcję energii. Gdy ten kontekst jest jasny, dużo łatwiej przejść do samego przebiegu reakcji i zobaczyć, gdzie dokładnie komórka inwestuje energię, a gdzie ją odzyskuje.
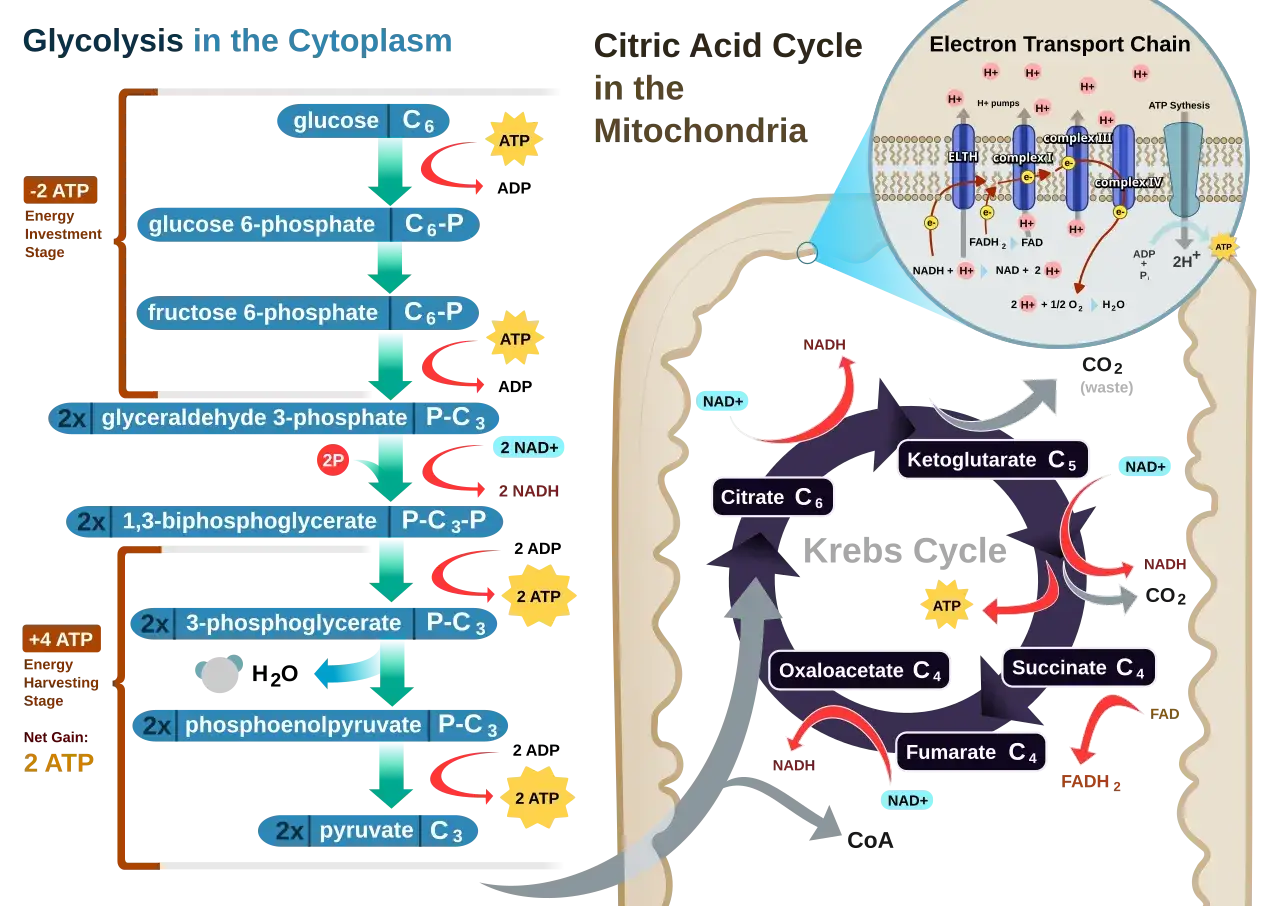
Jak przebiega rozkład glukozy krok po kroku
Cały szlak składa się z 10 reakcji enzymatycznych, ale do zapamiętania najwygodniej rozdzielić go na dwie fazy: inwestycji energii i zysku energetycznego. W pierwszej komórka zużywa ATP, żeby aktywować glukozę i przygotować ją do rozszczepienia. W drugiej odzyskuje energię w postaci ATP i NADH, a końcowym produktem są dwie cząsteczki pirogronianu.
| Faza | Co się dzieje | Bilans energetyczny | Dlaczego to ma znaczenie |
|---|---|---|---|
| Inwestycja | Glukoza jest fosforylowana, a potem rozszczepiana na dwa związki trójwęglowe. | Zużycie 2 ATP | Komórka „zamyka” glukozę wewnątrz i przygotowuje ją do dalszych reakcji. |
| Zysk | Dwie cząsteczki trioz są utleniane do pirogronianu. | Powstają 4 ATP i 2 NADH | Bilans netto wynosi 2 ATP, a NADH może później zasilać dalsze etapy oddychania komórkowego. |
Jeśli spojrzeć na to bardziej technicznie, najważniejsze punkty są trzy: heksokinaza lub glukokinaza fosforyluje glukozę, fosfofruktokinaza-1 wyznacza tempo całego szlaku, a kinaza pirogronianowa kończy produkcję ATP. Reszta reakcji przygotowuje substrat do tych trzech kluczowych momentów. Dla osoby uczącej się biochemii to cenna wskazówka: nie warto zapamiętywać wszystkiego z jednakową wagą, bo w praktyce kilka enzymów naprawdę decyduje o całym przebiegu.
Na końcu dostajemy 2 pirogroniany, 2 ATP netto i 2 NADH z jednej cząsteczki glukozy. To dobry punkt odniesienia, bo właśnie od tego zależy, czy komórka pójdzie dalej drogą tlenową, czy będzie musiała uruchomić fermentację. I to prowadzi nas do kolejnego pytania: co dokładnie dzieje się z pirogronianem po zakończeniu tego etapu?
Co dzieje się z pirogronianem po zakończeniu procesu
Końcowy produkt nie ma jednego losu. Wszystko zależy od dostępności tlenu, typu tkanki i aktualnego zapotrzebowania energetycznego. W komórkach tlenowych pirogronian trafia do mitochondrium i po przekształceniu do acetylo-CoA zasila cykl Krebsa. W warunkach beztlenowych albo wtedy, gdy komórka potrzebuje natychmiast odtworzyć NAD+, pirogronian może zostać zredukowany do mleczanu.
| Warunki | Los pirogronianu | Po co to służy | Przykład |
|---|---|---|---|
| Tlenowe | Przekształcenie do acetylo-CoA i wejście do mitochondrium | Dalsze pozyskiwanie energii z dużą wydajnością | Większość komórek w spoczynku |
| Beztlenowe u zwierząt | Redukcja do mleczanu | Odtworzenie NAD+, żeby szlak mógł trwać dalej | Mięśnie podczas intensywnego wysiłku, erytrocyty |
| Beztlenowe u drożdży | Fermentacja alkoholowa | Regeneracja NAD+ i utrzymanie produkcji ATP | Drożdże i część mikroorganizmów |
To właśnie regeneracja NAD+ jest tu krytyczna. Bez niej cały szlak zwalnia albo się zatrzymuje, bo jedna z reakcji utleniania nie ma już czego redukować. W komórkach bez mitochondriów, takich jak erytrocyty, to nie jest tylko „wariant awaryjny”, ale jedyny dostępny sposób na wytwarzanie ATP. Kiedy ta zależność jest jasna, sens regulacji całego procesu staje się znacznie bardziej czytelny.
Jak komórka reguluje tempo tego szlaku
Tempo nie jest stałe. Komórka reguluje je bardzo precyzyjnie, reagując na własny stan energetyczny, dostępność substratu i sygnały hormonalne. Najprościej mówiąc: jeśli ATP jest pod dostatkiem, szlak zwalnia; jeśli komórka ma deficyt energii, przyspiesza. To nie jest prosty włącznik i wyłącznik, tylko system wielopoziomowej kontroli.
| Enzym | Co go pobudza lub hamuje | Dlaczego jest ważny |
|---|---|---|
| Heksokinaza / glukokinaza | Heksokinaza jest hamowana przez glukozo-6-fosforan; glukokinaza w wątrobie reaguje na wyższą dostępność glukozy. | Kontroluje wejście glukozy do szlaku i jej „uwięzienie” w komórce. |
| Fosfofruktokinaza-1 | Aktywują ją AMP i fruktozo-2,6-bisfosforan, a hamują ATP i cytrynian. | To najważniejszy punkt kontroli tempa całego procesu. |
| Kinaza pirogronianowa | Aktywuje ją fruktozo-1,6-bisfosforan, a hamują ATP i alanina; w wątrobie wpływają na nią także insulina i glukagon. | Decyduje o końcowym etapie wytwarzania ATP i o tym, czy przepływ przez szlak utrzyma się na wysokim poziomie. |
W praktyce oznacza to, że komórka nie marnuje glukozy wtedy, gdy energii nie potrzebuje, i potrafi przyspieszyć przetwarzanie substratu, gdy warunki tego wymagają. Termin „allosteryczna regulacja” oznacza po prostu, że cząsteczki sygnałowe wiążą się z enzymem poza centrum aktywnym i zmieniają jego aktywność. To bardzo elegancki mechanizm, bo pozwala sterować szlakiem bez gwałtownych skoków. Z takiej perspektywy łatwo już przejść do tego, gdzie ten proces ma największe znaczenie w organizmie.
Dlaczego ten proces ma tak duże znaczenie w różnych tkankach
Najbardziej oczywisty przykład to mięśnie. Podczas intensywnego wysiłku potrzebują ATP szybko, a nie idealnie wydajnie, więc szlak glikolityczny staje się dla nich ważnym źródłem energii. Właśnie dlatego w sprintach, podnoszeniu ciężarów czy krótkich, bardzo intensywnych wysiłkach komórki mięśniowe mocno polegają na szybkim rozkładzie glukozy.
Drugi przykład to erytrocyty. Nie mają mitochondriów, więc nie mogą utleniać pirogronianu w klasyczny sposób. Dla nich ten szlak nie jest dodatkiem, ale podstawowym źródłem ATP. Podobnie ważny jest dla mózgu, który ma bardzo wysokie zapotrzebowanie na glukozę i źle znosi długotrwały niedobór energii. Wątroba z kolei pełni rolę regulatora: decyduje, czy glukoza ma zostać zużyta, zmagazynowana czy uwolniona do krwi.
- Mięśnie korzystają z tego szlaku, gdy potrzebują energii natychmiast, a tlen nie nadąża za zapotrzebowaniem.
- Erytrocyty są od niego całkowicie zależne, bo nie mają mitochondriów.
- Mózg zużywa glukozę bardzo intensywnie, więc sprawny przepływ przez ten szlak ma realne znaczenie dla funkcjonowania układu nerwowego.
- Wątroba pomaga utrzymać równowagę między zużyciem glukozy a jej magazynowaniem.
Warto też pamiętać, że w części komórek nowotworowych obserwuje się wysoki przepływ przez ten szlak nawet przy dostępie tlenu. To zjawisko nazywa się efektem Warburga i jest jednym z powodów, dla których metabolizm glukozy tak często wraca w biologii i medycynie. Nie oznacza to jednak, że każda komórka zachowuje się identycznie; kontekst tkankowy nadal ma znaczenie. Po takim ujęciu zostaje już tylko uporządkować najważniejsze fakty do zapamiętania.
Glikoliza bez wkuwania całej biochemii
Jeśli mam zostawić jedną prostą mapę do nauki, to wygląda ona tak: najpierw komórka inwestuje 2 ATP, potem rozszczepia glukozę na dwa związki trójwęglowe, a na końcu odzyskuje energię w postaci 4 ATP i 2 NADH. Taka kolejność pomaga zrozumieć nie tylko bilans, ale też logikę całego szlaku.
- Start: glukoza trafia do cytozolu i zostaje aktywowana przez fosforylację.
- Najważniejszy punkt kontroli: fosfofruktokinaza-1 decyduje o tempie przepływu.
- Bilans netto: 2 ATP z jednej cząsteczki glukozy.
- Produkty końcowe: 2 pirogroniany i 2 NADH.
- Los końcowy: zależy od tlenu, typu tkanki i dostępności mitochondriów.
Tak właśnie porządkuję ten temat, kiedy uczę go studentów albo sam wracam do biochemii po przerwie. Gdy zapamiętasz ten układ, dużo łatwiej zrozumiesz nie tylko sam proces, ale też jego związek z oddychaniem komórkowym, fermentacją i regulacją metabolizmu. To wystarczy, żeby ten fragment biochemii przestał być zbiorem wyjątków, a zaczął działać jak logiczna całość.