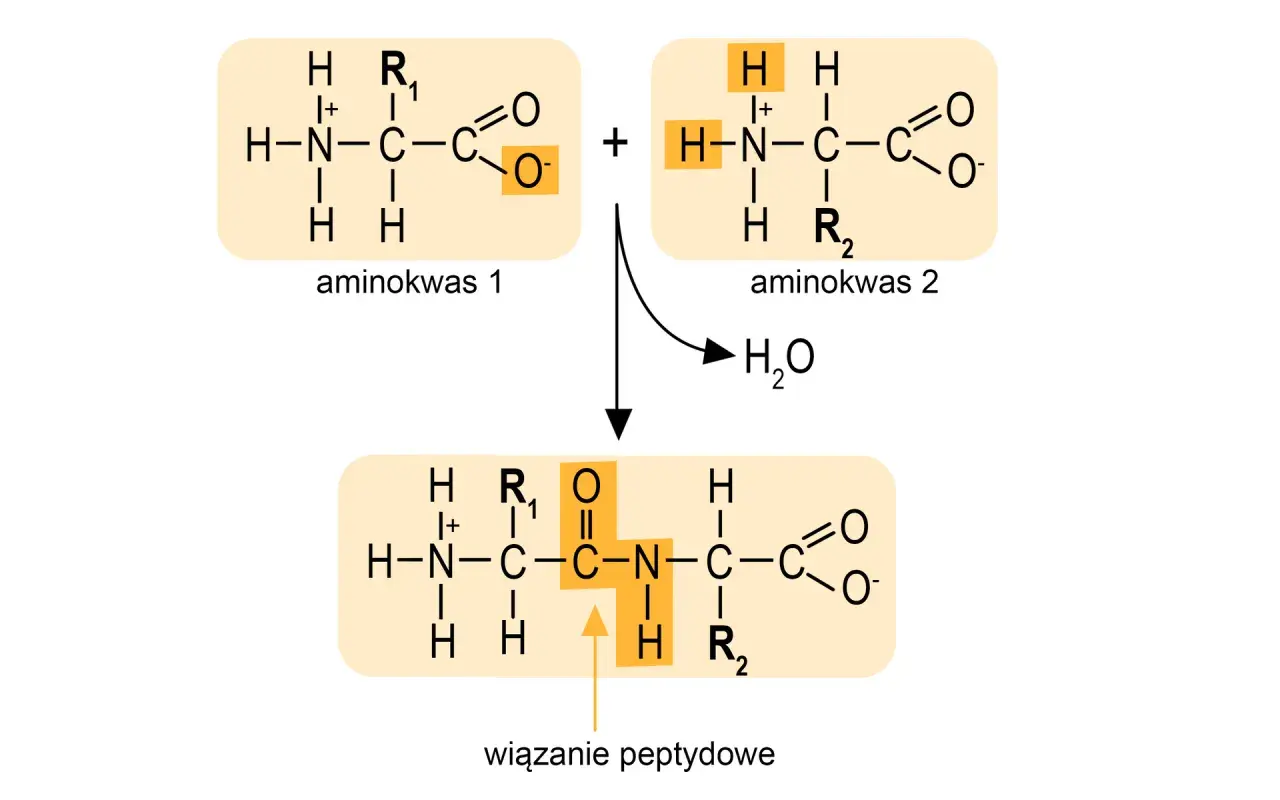
Najważniejsze fakty o tym połączeniu w białkach
- Powstaje między grupą karboksylową jednego aminokwasu a grupą aminową drugiego, z odłączeniem cząsteczki wody.
- Ma częściowy charakter podwójny, dlatego łańcuch białkowy jest planarny i ma ograniczoną rotację.
- W większości przypadków układ przyjmuje formę trans, a prolinę trzeba traktować jako ważny wyjątek.
- To właśnie ten fragment szkieletu białka w dużej mierze decyduje o fałdowaniu i stabilności struktur drugorzędowych.
- W zadaniach warto odróżniać go od zwykłego amidu i od połączeń izopeptydowych.
- Najwięcej błędów bierze się z mylenia chemii powstawania, geometrii i hydrolizy tego wiązania.
Jak powstaje i co dokładnie łączy
To połączenie tworzy się wtedy, gdy grupa karboksylowa jednego aminokwasu reaguje z grupą aminową drugiego. W uproszczeniu odłącza się cząsteczka wody, więc mówimy o reakcji kondensacji, a w biologii o syntezie prowadzącej do dipeptydu, tripeptydu albo dłuższego łańcucha. Po takim połączeniu mówimy już o resztach aminokwasowych, a nie o wolnych aminokwasach.
W komórce ten proces nie zachodzi spontanicznie w dowolnym tempie. Prowadzi go rybosom, a dokładniej centrum peptydylotransferazowe, które ustawia substraty we właściwej pozycji. W laboratorium chemicznym robi się to inaczej: aktywuje się grupę karboksylową, bo bez tego bezpośrednie łączenie aminokwasów byłoby zbyt mało wydajne.
To ważne rozróżnienie, bo student często widzi tylko efekt końcowy, a gubi mechanikę reakcji. Jeśli jednak w głowie zostanie prosty schemat: karboksyl + amina - woda = łańcuch, dalej cała reszta układa się znacznie logiczniej. Gdy ten etap jest jasny, można przejść do pytania, dlaczego ten fragment zachowuje się tak inaczej niż zwykłe wiązanie pojedyncze.
Dlaczego ten fragment łańcucha jest tak sztywny
Planarna geometria
Na zajęciach widzę, że najwięcej kłopotów bierze się z przekonania, że atomy C i N w tym układzie obracają się swobodnie jak w zwykłym wiązaniu pojedynczym. W rzeczywistości elektrony są zdelokalizowane, więc C-N ma częściowy charakter podwójny. To skraca długość tego połączenia do około 1,32 Å i wyraźnie ogranicza obrót wokół osi wiązania. Bariera rotacji jest duża, rzędu 20 kcal/mol, czyli około 80 kJ/mol.
W praktyce oznacza to, że szkielet białka nie jest elastycznym sznurkiem, tylko uporządkowaną konstrukcją. Taki układ jest stabilniejszy i łatwiej przewidzieć jego zachowanie w modelach przestrzennych. To dlatego w chemii białek tak często mówi się o planarnym fragmencie amidowym, a nie o zwykłym swobodnym połączeniu dwóch atomów.
Przeczytaj również: Co to jest mol w chemii i dlaczego jest tak ważny w nauce?
Układ trans i cis
Najczęściej sąsiednie fragmenty ustawiają się w formie trans, bo sterycznie jest ona korzystniejsza. Układ cis pojawia się rzadziej, ale przy prolinie staje się wyraźnie bardziej prawdopodobny, ponieważ jej pierścień ogranicza geometrię łańcucha. To właśnie dlatego proline bywa źródłem zaskoczeń w zadaniach i w analizie struktur.
Ten szczegół nie jest akademicką ciekawostką. Cis-trans wpływa na lokalny kształt łańcucha, a więc również na to, czy białko złoży się szybko, wolno, czy w ogóle przejdzie do aktywnej formy. Jeśli ktoś zna tylko definicję, zwykle nie rozumie jeszcze, skąd bierze się ta różnica w zachowaniu. Gdy zna geometrię, cała reszta zaczyna mieć sens.
Jak wpływa na budowę i fałdowanie białek
To właśnie ten sztywny fragment ustawia dalszą architekturę łańcucha. Grupy karbonylowe i amidowe szkieletu stają się odpowiednio akceptorami i donorami wiązań wodorowych, więc z tego samego tworzywa można zbudować helisę alfa, harmonijkę beta albo ostre zakręty w pętli. Gdyby obrót był całkiem swobodny, białka nie tworzyłyby tak stabilnych i powtarzalnych motywów.- Helisa alfa korzysta z regularnych wiązań wodorowych między kolejnymi odcinkami szkieletu.
- Harmonijka beta wymaga innego ustawienia fragmentów łańcucha, ale nadal opiera się na tej samej geometrii amidowej.
- Zakręty i pętle wykorzystują lokalne odchylenia od regularności i często pojawiają się tam, gdzie łańcuch zmienia kierunek.
- Prolina potrafi przerwać helisę albo wymusić ostrzejszy zakręt, bo jej budowa jest zbyt sztywna, by zachowywała się jak klasyczny aminokwas.
W fałdowaniu znaczenie ma też izomeryzacja cis-trans. Niektóre białka muszą przejść przez zmianę konformacji w tym miejscu, zanim osiągną aktywny kształt. To jeden z powodów, dla których w biologii strukturalnej tak dużą wagę przykłada się nie tylko do składu aminokwasowego, ale też do lokalnej geometrii całego szkieletu. Po takim spojrzeniu naturalnie pojawia się pytanie: czym to połączenie różni się od podobnych wiązań, które też występują w chemii organicznej i w białkach?
Czym różni się od innych podobnych połączeń
Najkrócej: każdy taki mostek w peptydzie jest amidowy, ale nie każdy amid jest peptydem. W praktyce liczy się to, jakie grupy reagowały i czy wiązanie leży w szkielecie łańcucha aminokwasów, czy poza nim. To rozróżnienie przydaje się zwłaszcza wtedy, gdy opis struktury jest skrócony albo gdy w zadaniu pojawiają się nietypowe połączenia sieciujące białka.
| Cecha | Mostek amidowy w peptydzie | Połączenie izopeptydowe | Zwykły amid |
|---|---|---|---|
| Gdzie powstaje | Między grupą karboksylową i aminową szkieletu aminokwasów | Przez grupy boczne albo między grupą boczną a szkieletem | W dowolnych związkach organicznych zawierających układ amidowy |
| Co łączy | Reszty aminokwasowe w głównym łańcuchu | Fragmenty boczne, czasem sieciujące całe białka | Dowolne fragmenty organiczne |
| Najważniejsza cecha | Planarność i ograniczona rotacja | Może zmieniać architekturę białka poza szkieletem | Nie musi mieć znaczenia biologicznego |
| Pułapka w zadaniach | Łatwo pomylić je ze zwykłym wiązaniem pojedynczym | Bywa mylone z głównym szkieletem białka | Nie każdy amid uczestniczy w budowie peptydów |
Drugie częste nieporozumienie dotyczy trwałości. To połączenie jest stabilne w warunkach fizjologicznych, ale może ulec hydrolizie w silnie kwaśnym lub zasadowym środowisku, a także pod wpływem enzymów proteolitycznych. Warto więc odróżniać chemiczną odporność fragmentu od biologicznego rozcinania go przez enzymy, bo to nie jest to samo. Gdy to rozróżnienie jest jasne, można przejść do samego odczytywania zapisów i modeli.
Jak czytać zapis łańcucha i nie pomylić kierunku
W zapisie liniowym patrzę najpierw na dwa końce łańcucha: N-koniec ma wolną grupę aminową, a C-koniec wolną grupę karboksylową. Sekwencję zapisuje się zwykle od N do C, więc kierunek ma znaczenie nie tylko w biologii, ale też w chemii i bioinformatyce. Jeśli w zadaniu widzisz skrót aminokwasów, najpierw ustal właśnie ten kierunek.
- Znajdź w szkielecie atom węgla karbonylowego i sąsiedni atom azotu.
- Sprawdź, czy łańcuch jest prowadzony od N-końca do C-końca.
- Oceń, czy fragment ma układ trans czy cis, jeśli masz model 3D albo dane konformacyjne.
- Zwróć uwagę, czy połączenie nie przebiega przez grupę boczną, bo wtedy możesz mieć do czynienia z połączeniem izopeptydowym.
Na modelach kulkowo-kreskowych planarny charakter tej grupy zwykle widać bardzo dobrze, ale w zwykłym wzorze kreskowym łatwo go przegapić. Dlatego przy analizie warto wyrobić sobie prosty nawyk: najpierw szkielet, potem łańcuch boczny, dopiero na końcu szczegóły stereochemiczne. To oszczędza sporo pomyłek, zwłaszcza przy większych peptydach.
Trzy reguły, które oszczędzają najwięcej błędów
- Nie traktuj C-N jak zwykłego, swobodnie obracającego się połączenia pojedynczego.
- Zawsze sprawdzaj układ trans/cis, zwłaszcza gdy w sekwencji pojawia się prolina.
- Rozróżniaj powstawanie łańcucha, jego geometrię i hydrolizę, bo to trzy różne poziomy opisu.
Jeśli mam zostawić jedną myśl końcową, to taką: ten fragment szkieletu decyduje o tym, czy aminokwasy pozostaną tylko listą cegiełek, czy złożą się w działające białko. Kiedy zaczyna się patrzeć na łańcuch przez pryzmat rezonansu, ograniczonej rotacji i lokalnej geometrii, biochemia staje się dużo bardziej logiczna, a zadania z chemii przestają wyglądać na zlepione z przypadkowych faktów.